 English
English Español
Español Français
Français Deutsch
Deutsch Italiano
Italiano Português
Português
L’expérience réalisée par l’équipe de Monod1, a soulevé un paradoxe : la bactérie femelle transformée par l’entrée d’un chromosome mâle conserve le phénotype « constitutif » pendant 90 minutes, puis manifeste le phénotype « inductible ». Les bactéries sur lesquelles Monod avait travaillé au début de ses recherches sont dites inductibles au sens où elles ne fabriquent l’enzyme qui assure l’hydrolyse du lactose que lorsque celui-ci est placé dans le milieu nutritif. Le lactose induit la fabrication de l’enzyme, c’est le mécanisme de cette induction que l’on recherche.
La mutation conférant le phénotype « constitutif » entraine la perte de l’inductibilité, savoir la fabrication de l’enzyme même en absence de lactose. C’est par la sélection de diverses souches et par des expériences de conjugaison entre bactéries, laquelle équivaut à des croisements reproductifs, que Monod et son équipe ont avancé dans l’élucidation du mécanisme de l’induction. Ainsi ils réalisent l’expérience dite « spaghetti » où le chromosome mâle pénètre petit à petit, si bien que tous les gènes qu’il porte ne s’expriment pas en même temps. Ils sont amenés à postuler l’existence d’un gène d’inductibilité noté I et admettant 2 allèles : I+ et I-.
Pour expliquer le délai de 90 minutes avant que la bactérie femelle hybride, constitutive au début, soit devenue inductible, il faut imaginer que le gène I est situé en queue de chromosome, entrant ainsi tardivement dans la bactérie femelle. C’est le deuxième niveau d’explication du paradoxe1. Le troisième niveau d’explication est entrevu par François Jacob, frappé soudainement, à l’été 1958, par la similitude entre ses propres expériences « spaghetti » menées sur les bactériophages quelques années plus tôt3, et les expériences menées conjointement avec Monod sur les bactéries inductibles et constitutives. Il écrira :
Et soudain, un éclair. L’éblouissement de l’évidence. Comment ne pas y avoir pensé plus tôt ? (…) Dans les deux cas, un gène gouverne la formation d’un produit cytoplasmique, un répresseur qui bloque l’expression d’autres gènes, empêchant ainsi soit la synthèse de la galactosidase, soit la multiplication des virus.
(cité par Debré p. 218)
Quand le « nouveau converti » résiste
L’allèle I+ doit donc coder pour une protéine qui agit comme répresseur de l’allèle Z+ , celui de la β-G-ase. Lorsque le gène I a pénétré au bout des 90 minutes, il entraine la fabrication de la protéine qui empêche Z+ de s’exprimer, et voilà comment la bactérie est devenue inductible, l’état ordinaire sauvage. Mais comment agit le lactose au titre d’inducteur de la β-G-ase ? Est-ce qu’il empêche le répresseur d’être fabriqué sous le contrôle de I+ ou est-ce qu’il intervient sur la protéine répresseur ? Dans la première hypothèse, cela voudrait dire qu’il agirait directement sur le gène I. Mais n’allons pas si vite, car Monod n’est pas convaincu.
Pour rappel, Jacob étudiait le phénomène de lysogénie3, cas de bactéries qui hébergent des virus (nommés phages) sans dommage pour elles.
Il aboutit à la conclusion que le phénomène est dû à ce que le phage est en quelque sorte « endormi », on le nomme prophage. Autrement dit, l’ADN du prophage est lui aussi à l’état réprimé, son expression est bloquée. C’est ainsi qu’il fait le rapprochement avec l’induction : chez Escherichia coli, l’allèle de la β-G-ase est en temps normal réprimé, bloqué ; et c’est le lactose qui fait lever cette répression.
Or Monod résiste à cette interprétation. « Converti » pourrait-on dire entretemps au darwinisme, il craint un retour au néolamarckisme, car l’hypothèse de Jacob revient à redonner un rôle à l’environnement, représenté ici par le lactose jouant ce rôle d’inducteur, dans l’expression d’un gène. On se souvient de son combat contre le savant soviétique Lyssenko4, qui a sûrement contribué à sa conversion au darwinisme, en même temps que l’avancée opérée par la génétique achevait de le convaincre.
C’est en effet dans les années 1930-1940 qu’est élaborée la théorie synthétique de l’évolution dans laquelle la variabilité du génome donne prise à la sélection naturelle. Tandis que dans le dogme néo lamarckien, les caractères acquis sous l’influence du milieu pouvaient être transmis à la descendance. Du reste Darwin lui-même, qui ne connaissait pas la génétique, admettait ce postulat. Dans le nouveau dogme de la génétique, le gène est devenu tout-puissant, il est souverain, il ne saurait recevoir d’ordres de l’extérieur.
La logique du double mécanisme de répression / levée de la répression, la seconde sous l’action d’un facteur externe, passe mal l’interdit de la génétique. Néanmoins, après une série de discussions entre les deux hommes, Monod accepte l’explication de Jacob. Nous sommes à l’automne de 1958. C’est à partir de ce moment que Monod, comme ce fut le cas antérieurement, va s’employer à compléter l’explication de Jacob pour finir par élaborer le modèle définitif de l’opéron lactose.
On aura noté combien souvent Monod a résisté, tout au long de ce long chemin vers la découverte qui lui a valu le prix Nobel, avec Lwoff et Jacob.
Il a hésité à abandonner la théorie en vigueur au début des années 1950 de l’adaptation enzymatique, malgré la diauxie (voir ici Quand Jacques Monod passe de la biométrie à la génétique) et d’autres résultats expérimentaux en sa défaveur. Il n’a pas tout de suite compris l’intérêt de la découverte de Beadle et Tatum et leur formule restée célèbre « un gène une protéine ». Il n’a pas été convaincu immédiatement par ses collaborateurs quant à la perméabilité sélective. Il n’admet pas tout de suite l’hypothèse sur la répression que l’américain Szilard lui sert pourtant sur un plateau2. Et c’est Jacob qui a « l’insight » du lien entre l’affaire du lactose et celle de la lysogénie !
Mais ce sont ses objections et ses réticences qui forcent à trouver des démonstrations expérimentales plus éclatantes. Et c’est lui qui dirige les équipes. Enfin c’est lui qui opère les synthèses, bâtit et améliore le modèle.
Son mérite est donc intact, mais là n’est pas le lieu d’un quelconque jugement.
Epilogue de 25 ans de recherches : l’opéron lactose
À cet automne 1958, il est bien établi :
que l’induction de la bêta-galactosidase par le lactose relève d’un mécanisme de levée d’inhibition de l’expression du gène de cette enzyme.
et que c’est une protéine sous le contrôle d’un autre gène qui empêche justement certains gènes de s’exprimer, autrement dit de diriger la fabrication des protéines correspondantes.
Mais le rôle de l’inducteur n’est pas encore bien clair. Les travaux vont donc se poursuivre jusqu’à établir le schéma complet. Ainsi, on identifie une nouvelle mutation au phénotype constitutif qui fournit un résultat inattendu aux expériences de croisement.
En effet, croisée à la souche sauvage inductible classique, cette bactérie reste constitutive, c’est-à-dire que, aussi bien en absence qu’en présence de l’inducteur, elle fabrique la β-Gase. Il faut alors admettre que l'allèle muté se comporte comme s’il était dominant par rapport au sauvage, ce qui est très inhabituel en génétique classique.
L’interprétation de ce comportement étrange est que l’on n’a pas affaire à un gène au sens classique, codant pour une enzyme fonctionnelle, mais à une séquence particulière d’ADN qui sera baptisée site opérateur (noté O), contrôlant la synthèse d’une protéine-opérateur.
Le schéma de fonctionnement de l'opéron serait alors le suivant : en absence d'inducteur, le répresseur fabriqué sous contrôle du gène I reconnaît spécifiquement le site O et s'y fixe. L'encombrement de ce complexe est tel qu’il bloque l’expression de l'ensemble des gènes de la série.
Fig.1 – Schéma de l’opéron lactose (avant-dernière version)
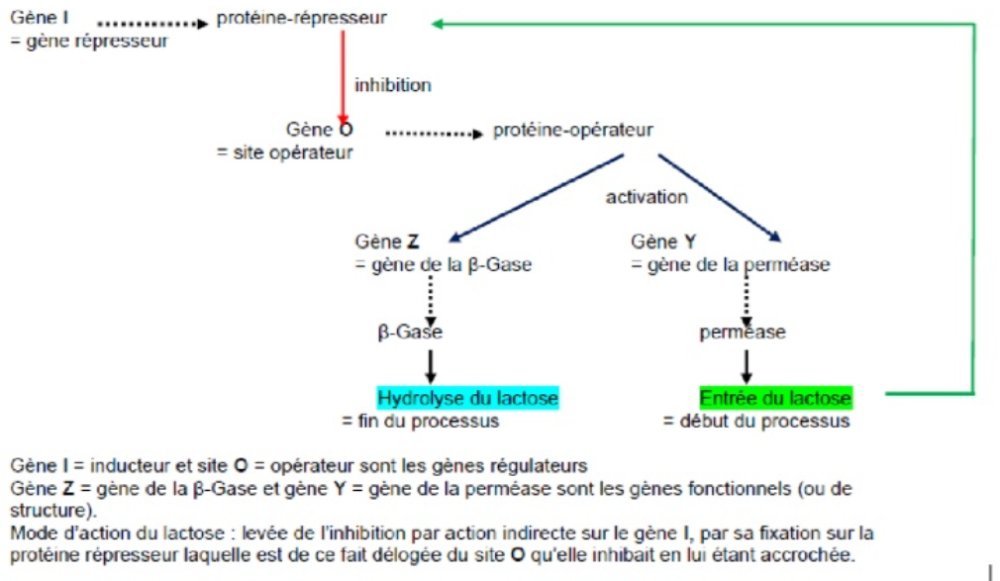
Monod suggère qu’il doit exister une souche mutante de bactérie au niveau du gène O qui ne réponde plus au répresseur. Jacob se souvient qu’il a déjà trouvé un phage mutant qui a les mêmes propriétés. La mutation recherchée est obtenue : cette souche a le phénotype constitutif puisque l’allèle Oc (O exposant c pour constitutif) par quoi on désigne la mutation, ne peut pas fixer la protéine répresseur.
C’est en 1966 que des chercheurs américains (Hill et Gilbert) auront isolé le répresseur et confirmé sa nature protéique, ce dont Monod n’avait jamais douté, et tandis qu’il s’était déjà tourné vers d’autres sujets de recherches, et bien d’autres activités présentées au tout début de cette série d’articles (voir ici même Monod, un biologiste dans le siècle).
La découverte de Monod est-elle restée sans suites, ou bien a-t-elle permis d’identifier d’autres opérons ? Et vers quoi l’existence de mécanismes de régulation de ce type a-t-elle engagé la biologie ? Quelle aura été la portée de la découverte de Lwoff, Monod et Jacob ? Et à quoi s’est alors consacré Jacques Monod après avoir établi le schéma définitif de l’opéron lactose (voir Fig. 2) ?
On notera que la figure 2 mentionne le rôle de l’ARN messager, non évoqué jusqu’ici, et pour cause : c’est après avoir établi l’avant-dernière version de l’opéron lactose que Monod et son équipe vont se pencher sur un épineux problème jusqu’ici peu abordé : comment se fait l’expression d’un gène lorsqu’il n’est pas réprimé ? Autrement dit quelle est la séquence des événements qui mènent du gène à la protéine dont il contrôle la fabrication ?
Fig. 2 Le schéma définitif de l’opéron lactose
2-a En absence de lactose.
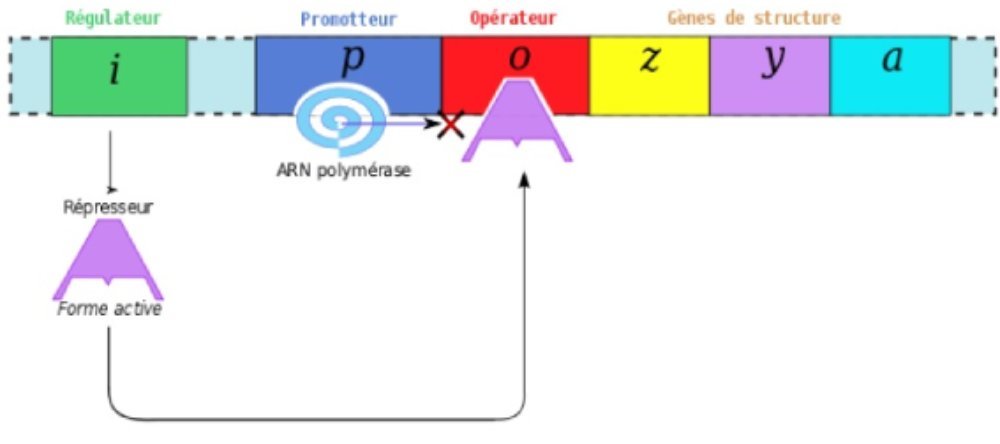
En absence de lactose : le répresseur est fixé sur le gène opérateur, ce qui empêche l’action de l’ARN polymérase, donc de la transcription des gènes dont celui de la β-Gase.
2-b En présence de lactose.
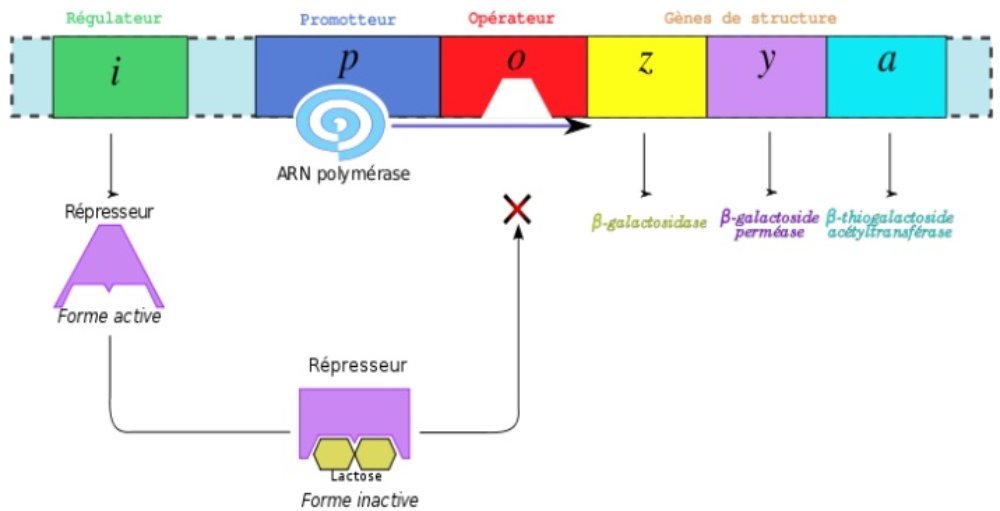
En présence de lactose : en se fixant sur le répresseur, le lactose ne permet plus à ce dernier de se fixer sur le gène opérateur, et l’ARN polymérase peut agir pour que soit transcrit l’ADN en ARN messager, condition à son tour de la synthèse protéique.
Bibliographie
Debré, Patrice, (1996), Jacques Monod, Flammarion.
1 Monod 14, le schéma de l’induction se précise dans Meer.
2 Monod 13, induction vs constitution dans Meer.
3 Monod 12, un virus au service de l’induction enzymatique dans Meer.
4 L’affaire Lyssenko, combat mémorable de Jacques Monod dans Meer














