English
English Español
Español Français
Français Deutsch
Deutsch Italiano
Italiano Português
Português
Monod et Jacob se sont mis à travailler ensemble à partir de 1957, tirant profit d’expériences de croisements entre souches bactériennes, puisque la sexualité des bactéries a été découverte quelques années plus tôt aux USA. On peut ainsi faire de la génétique classique en croisant des souches divergeant par divers caractères isolés à partir de mutations, comme l’avaient fait Mendel au siècle précédent avec ses petits pois puis Morgan avec ses mouches drosophiles dans la première moitié du XXe siècle. Il s’agit de l’expérience dite « spaghetti » (voir ici Monod 12 Un virus au service de l’induction1). La technique est perfectionnée au point qu’on peut interrompre l’entrée du chromosome mâle à différents moments, ce qui permet de localiser les gènes les uns par rapport aux autres, à l’instar des cartes génétiques qu’avait établies Morgan.
Les conditions techniques sont donc réunies pour que Monod et Jacob avancent dans l’élucidation du mécanisme de l’induction enzymatique impliquant des gènes. Or, à ce stade, ils sont un peu bloqués au plan conceptuel. Deux gènes ont été mis en évidence : celui qui dirige la synthèse de la galactose-perméase, laquelle contrôle l’entrée du lactose au niveau de la membrane ; le gène de la bêta-galactosidase, laquelle enzyme « casse » la molécule de lactose. Mais comment expliquer que la présence de lactose dans le milieu de culture induise la fabrication de la β-G-ase, laissant entendre que son gène serait inactif en l’absence de lactose ? Qu’est-ce qui rendrait ce gène inactif ? Où et comment agirait le lactose ? Et en quoi la perméase membranaire est-elle concernée ?
En 1958, un biochimiste américain, Arthur Pardee, arrive à l’Institut Pasteur pour y passer une année. Grâce à lui, les techniques de mesure pour obtenir les courbes de croissance vont être simplifiées, ce qui va donner un coup d’accélérateur aux recherches. La suite du travail sera appelée « expérience pyjama ». Les résultats sont publiés dans le Journal of Molecular Biology, article resté très célèbre sous la première dénomination PA-JA-MO rapidement convertie, par boutade, en PY-JA-MA (The genetic control and cytoplasmic expression of « inducibility » in the synthesis of β-galactosidase by Escherichia coli, by A .B. Pardee, F. Jacob and J. Monod, J. Mol. Biol.,1959, 1, 165-178).
« Pyjama » est un jeu de mots issu de « pajamo », formé par les premières lettres des trois noms Pardee/Jacob/Monod.
Première étape de l’expérience pyjama
Si la galactose-perméase et la bêta-galactosidase sont toutes deux nécessaires à la consommation du lactose par la bactérie, l’une pour permettre l’entrée du lactose, la seconde pour l’hydrolyser, aucun lien direct entre ces deux enzymes n’avait été établi.
Or voici qu’on isole deux souches de bactéries aux curieuses propriétés. L’une ne produit de façon coordonnée la perméase et la β-G-ase que sous l’induction du lactose. Cette souche est qualifiée d’inductible. En ce sens qu’elle ne fabrique pas spontanément et d’emblée les deux enzymes, mais seulement lorsque le lactose est introduit dans le milieu de culture. Le mystère s’épaissit : que le lactose induise la production de β-G-ase, soit ; mais qu’il induise en outre la fabrication de la protéine membranaire qui permet son entrée même dans la cellule bactérienne, voilà qui est bien étrange !
L’autre souche utilisée est appelée « constitutive ». Elle produit spontanément, et d’emblée à taux maximal, les deux enzymes, qu’il y ait ou non du lactose dans le milieu ; autrement dit elle ne présente pas le phénomène d’induction. L’existence de ces deux souches confirme, s’il en était besoin à ce stade des travaux de Monod, l’intervention des gènes. Les deux souches possèdent évidemment le gène de la β-G-ase (noté Z) et celui de la galactose-perméase (noté Y). Mais comment se fait-il que chez l’une, la souche « constitutive », aucune induction ne se manifeste ?
Doit-on admettre l’existence d’un gène qui serait impliqué dans le phénomène d’induction, et qui chez la bactérie « constitutive » devrait être altéré par la mutation ? Ce n’est pas encore, à ce stade, l’intuition de Monod et Jacob. Ce qui ne les empêche pas de faire de la génétique classique avec leurs bactéries. Par la sélection des souches mutantes, ils ont obtenu une souche « constitutive » particulière, dépourvue du gène de la β-G-ase. Pour respecter la terminologie des généticiens et surtout pour mieux comprendre l’expérience, il est préférable de parler de l’allèle de la β-G-ase : un gène peut en effet présenter des variantes (en général issues de mutations) ; on dit qu’à l’emplacement donné d’un gène peuvent se trouver diverses variantes ou allèles de celui-ci.
En l’occurrence ici, l’une des souches possède l’allèle de la β-G-ase, qui est noté Z+. L’autre contient un allèle muté de ce gène, noté Z-, si bien qu’elle est incapable de fabriquer la β-G-ase, même en présence de lactose. Voici l’expérience. Dans un milieu dépourvu de l’inducteur (autrement dit sans lactose), les chercheurs croisent une bactérie mâle avec une bactérie femelle dont les caractères sont les suivants.

Tableau 1 – Croisement de souches inductibles et constitutives particulières.
Rappelons que la souche constitutive d’origine produit d’emblée la perméase et la galactosidase. Celle utilisée ici ne produit pas la β-G-ase du fait de l’allèle Z-. Le résultat est donc paradoxal puisque le milieu est dépourvu de lactose (l’inducteur) et que le génotype de la bactérie femelle ne lui permet pas de fabriquer la β-G-ase. La conclusion s’impose : l’allèle Z+ de la β-G-ase a bien été transféré de la bactérie mâle inductible à la bactérie femelle qui en était dépourvue, par l’entrée du chromosome (ou d’un segment de celui-ci) de la bactérie mâle dans la bactérie femelle.
Mais quelle est l’origine de ce phénotype « constitutif » de la souche de bactérie ainsi nommée ? Pour Monod à ce moment, l’explication n’est toujours pas dans les gènes mais serait à rechercher plutôt dans l’existence d’un inducteur endogène, agissant comme le lactose mais interne à la bactérie. On se trouve à nouveau dans l’impasse, car comment identifier cet hypothétique inducteur endogène ?
Quand un physicien nucléaire s’en mêle
Outre Pardee, arrive aussi à l’Institut Pasteur Leo Szilard, physicien nucléaire américain d’origine hongroise. Il s’est reconverti dans la biologie après le désastre humain provoqué par l’explosion des bombes atomiques sur Hiroshima et Nagasaki au Japon à la fin de la Seconde Guerre mondiale.
Szilard propose à Monod une autre hypothèse pour expliquer, a contrario, l’inductibilité, au lieu de chercher l’explication de la constitutivité. Szilard imagine que c’est l’expression des gènes qui serait empêchée en temps normal et que cette répression des gènes serait levée par l’addition de l’inducteur. Jusqu’ici, pour expliquer l’absence d’expression d’un gène, la génétique estimait que c’était dû à la présence d’un allèle ne permettant pas le contrôle de la fabrication de la protéine. Comme ici l’allèle Z- .
Le schéma de ce mécanisme supposé de Szilard peut être représenté ainsi.
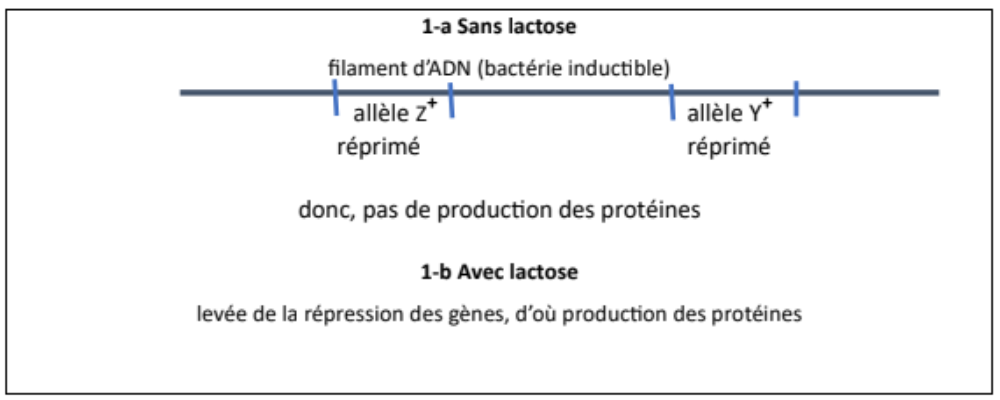
Figure 1 – Premier schéma de l’induction comme levée de la répression.
Insistons : ce sont bien les allèles Z+ et Y+ qui se trouvent sur le chromosome, ils ont la capacité de diriger la synthèse des protéines correspondantes, mais s’y trouvent empêchés ; on ne sait pas encore comment. Tout ce qui peut être dit à ce stade, c’est la conclusion apparente : l’inducteur lève la répression des gènes qui peuvent alors s’exprimer. La négation de la négation redonne bien l’affirmation, enfin plutôt ici l’induction. Complication excessive ou relance de dialectique hégélienne chère à Karl Marx, autant dire que Monod n’est pas convaincu. Jacob et lui ne s’avouent pourtant pas vaincus !
Notes
1 Monod 12, un virus au service de l’induction enzymatique.
Bibliographie
Debré, Patrice, (1996), Jacques Monod, Flammarion.