English
English Español
Español Français
Français Deutsch
Deutsch Italiano
Italiano Português
Português
Le problème posé par la découverte du phénomène d’induction a occupé Jacques Monod pendant de longues années. Pour rappel, chez la bactérie Escherichia coli dont il étudie le métabolisme nutritif, le lactose, sucre nourricier, a besoin d’être hydrolysé pour être catabolisé. Cette opération dépend d’une enzyme, la β-Galactosidase (en abrégé β-G-ase) qui n’apparait dans le cytoplasme cellulaire que s’il y a du lactose dans le milieu. Le lactose semble induire la présence dans la cellule bactérienne de l’enzyme même qui va le dégrader. C’est étrange, mais on dispose alors de la théorie de « l’adaptation enzymatique » qui postule que le substrat, en se fixant sur une protéine dite précurseur déjà présente, en modifie la structure tridimensionnelle, ce qui la rend active pour accomplir sa fonction (voir ici Jacques Monod : De l’adaptation à l’induction enzymatique).
Au début, Monod adhère à cette explication, dont le côté mécaniste, instructif peut-on dire lui convient en ce milieu du XXe siècle : le substrat instruit directement la protéine qui va catalyser sa transformation. La thèse sélective liée à la génétique n’a pas encore très bonne presse chez les scientifiques français, marqués par le néolamarckisme, et où elle reste de toute façon mal connue. Lamarck est un naturaliste français de la fin du XVIIIe siècle qui proposa une thèse évolutionniste précédant celle de Darwin mais dont les bases sont différentes, notamment l’absence de sélection. Elle dominera la biologie française sous l’influence de Claude Bernard, célèbre physiologiste français du XIXe siècle.
L’évolution de la pensée de Monod, décrite dans la série d’articles déjà consacrés (Monod 1 à Monod 13) prendra du temps, sous l’influence des conceptions qui changent, des connaissances qui se développent et aussi des collaborations et des rencontres qui l’ont jalonnée. Une affaire d’équipe, de discussions parfois serrées, de protocoles expérimentaux rigoureux, où Monod s’affirme comme le « patron » qui impulse les travaux et assure la communication, capable aussi de reconnaitre ses torts.
Encore un paradoxe
Parvenue à la conclusion de l’implication des gènes dans le phénomène d’induction, l’équipe de Monod peine encore à élucider le mécanisme menant des uns à l’autre. L’on vient de voir (Monod 13) que l’idée d’une répression des gènes comme état « normal » est due au physicien Léo Szilard reconverti dans la biologie. Or les expériences dites « pyjama » de croisements (par conjugaison) entre souches bactériennes distinctes ont montré simplement (si l’on peut dire !) qu’en passant d’une cellule (qualifiée de mâle) à l’autre (femelle) le chromosome entrant transforme la bactérie par la présence de ses allèles particuliers. Cela ne suffit pas pour établir le mécanisme de l’induction comme levée de la répression, selon la suggestion de Szilard.
Monod et Jacob décident alors de suivre la synthèse de la β-G-ase dans le temps, après la conjugaison bactérienne. En effet, la production de l’enzyme débute seulement 3 minutes après la mise en contact des bactéries mâle et femelle. En répétant leurs mesures à intervalles réguliers, les chercheurs observent que la production de β-G-ase se poursuit comme attendu d’une bactérie conjuguée de phénotype « constitutif », qui produit l’enzyme sans que le lactose soit présent dans le milieu. Mais ô surprise, cette production cesse au bout de 90 minutes, et reprend dès qu’on ajoute du lactose. Ce qui laisse supposer que la bactérie conjuguée est devenue, curieusement, « inductible ». Trois niveaux d’explications seront successivement proposés à ce paradoxe.
Premier niveau
La bactérie femelle utilisée dans cette conjugaison est « constitutive » = ne présente pas le phénomène d’induction au lactose. Mais il s’agit d’une souche particulière, qui n’est pas la « constitutive » d’origine (voir Monod 12) car elle est dépourvue de l’allèle Z+, qui contrôle la production de β-G-ase ; elle est Z-, (voir Monod 13). Après le croisement, elle acquiert Z+, apporté par la bactérie mâle de souche « inductible ». À preuve, 3 minutes après la conjugaison la β-G-ase est fabriquée, et ce durant 90 minutes, en milieu dépourvu de lactose. La bactérie conserve le phénotype « constitutif » pendant cette durée, mais elle est cette fois constitutive comme la souche d’origine (voir Monod 12) capable de fabriquer sans induction à la fois la β-G-ase et la perméase. Vous suivez toujours ? En fait ce premier niveau ne fait que proposer une analyse des résultats obtenus. Passons au niveau suivant.
Deuxième niveau
Au bout de 90 minutes, la bactérie hybride est devenue inductible. Comment son phénotype a-t-il pu changer alors que son génotype semble ne pas avoir changé ? Les chercheurs suspectent qu’autre chose que le chromosome mâle ait pu pénétrer dans la bactérie femelle. Or des expériences de contrôle excluent cette hypothèse. L’expérience « spaghetti » (voir Monod 13) avait mis en évidence une entrée progressive du chromosome mâle, à l’image du filament de pâte qu’on avale. L’idée émerge qu’un autre gène que les deux évoqués jusqu’ici puisse être impliqué dans le phénomène. Baptisé « gène d’inductibilité », il est noté I et admettrait 2 allèles : I+, et I-. Pour expliquer le délai de 90 minutes avant que la bactérie femelle hybride, constitutive au début, soit devenue inductible, il faut imaginer que le gène I est situé en queue de chromosome, entrant ainsi tardivement dans la bactérie femelle.
En outre, on doit admettre que l’allèle I+, (sauvage) de l’inductibilité domine l’allèle I-, (muté) de la constitutivité. Jusqu’ici tout va bien. Quoique : comment l’allèle I+, confère-t-il le phénotype inductible ? En vertu du principe « un gène une protéine », cet allèle contrôle indubitablement la fabrication d’une protéine : mais quelle serait la fonction de cette protéine ? Le niveau suivant va tenter de répondre.
Troisième niveau
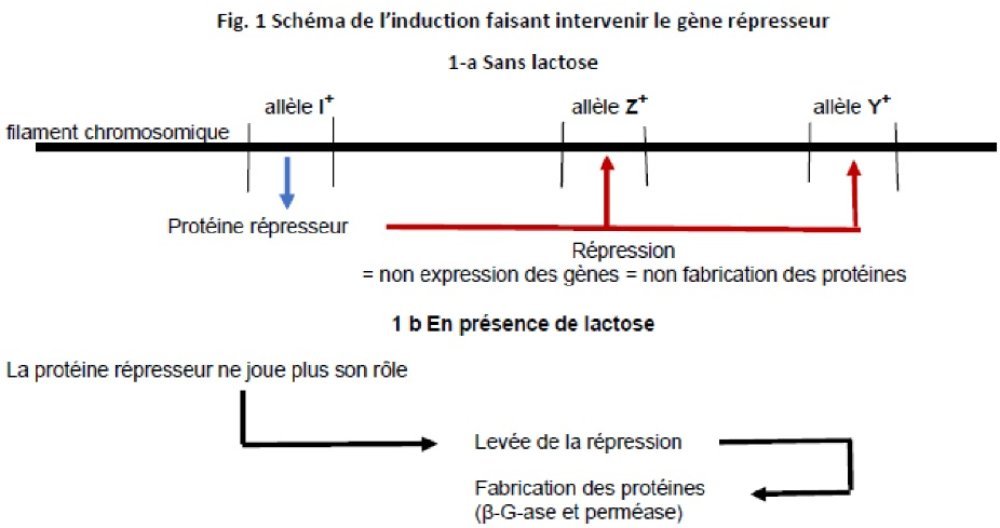
Il suffit d’admettre que l’allèle I+, code pour une protéine qui agit comme répresseur de l’allèle Z+, de la β-G-ase. Sans lactose dans le milieu, la bactérie inductible voit son allèle Z+, réprimé, empêché par la protéine répresseur codée par l’allèle I+. En présence de lactose, cette répression est levée, sans que l’on sache encore comment, et Z+, actif peut diriger la synthèse de l’enzyme.
Nouveau schéma du mécanisme de l’induction
Avec ce troisième niveau d’explication du paradoxe, Monod dresse un schéma plus complet et plus précis du mécanisme de l’induction (voir Fig. 1)
Ce schéma a permis de franchir un pas important. Il soulève néanmoins un autre problème : comment agit le lactose pour lever la répression ? Soit il agit directement sur le gène répresseur, soit il agit sur la protéine répresseur. Le chemin semble donc tracé. N’y a-t-il plus qu’à trouver le moyen de départager ces hypothèses ? Nous ne sommes pas au bout des surprises.
Bibliograhpie
Debré, Patrice, (1996), Jacques Monod, Flammarion.