English
English Español
Español Français
Français Deutsch
Deutsch Italiano
Italiano Português
Português
Qu’est-ce que le vivant ? La question n’en finit pas d’être posée et reposée, tant par les philosophes, les biologistes que le commun des mortels. Buffon, notre naturalise philosophe (voir dans cette revue Buffon le naturaliste philosophe), auteur d’une célébrissime Histoire naturelle, en signe paradoxalement la mort. L’histoire naturelle est en effet surtout descriptive alors qu’il voudrait fonder une science explicative du vivant, celle que Lamarck popularisera bientôt sous le nom de biologie. Pour cela, il faut s’attaquer au « noyau dur » de ce qui demeure le plus énigmatique dans le vivant : comment il est organisé, se nourrit et se reproduit.
Au cours du XVIIe siècle et au moins jusqu’au milieu du XVIIIe siècle, la controverse fait rage sur le problème de la génération comme on la nomme alors, savoir comment se forment puis se développent l’œuf et l’embryon. Ni compromis ni dépassement ne semblent en vue.
Épigenèse vs préformation
L’épigenèse considère que le germe du nouvel être – dit de façon actuelle, l’œuf ou l’embryon – construit progressivement ses organes, lesquels apparaissent au fur et à mesure avant de grandir. Présente déjà chez Aristote (-384-322), l’idée est soutenue en particulier par Descartes (1596-1650) pour des raisons théoriques qui veulent que le mouvement doit suffire à expliquer l’organisation spontanée de la matière. La préformation, présente chez Hippocrate au Ve siècle avant notre ère, est défendue par l’italien Marcello Malpighi (1628-1694) et reprise par le hollandais Jan Swammerdam (1637-1680). Cette thèse estime que le germe contient tous les organes en miniature, tellement petits qu’on ne peut pas les voir, et que le développement n’est que le déploiement, suivi de la croissance, de ces organes préformés. La préformation a le mérite de fournir une explication rationnelle à la formation des organes qui reste mystérieuse dans l’épigenèse. Elle a même une coloration étrangement mécaniste. En fait, ni l’une ni l’autre ne résolvent l’énigme de la transmission d’une génération à la suivante. C’est pourquoi certains « préformationnistes » imaginent que les germes préexistent de tout temps, ou depuis la création, et sont emboîtés les uns dans les autres pour toutes les générations à venir, à la manière des poupées russes : c’est la théorie de la préexistence des germes, défendue par Swammerdam. Raison probable pour laquelle Descartes est hostile à la préformation.
Cette controverse va s’affaiblir au milieu du XVIIIe siècle, lorsque Wolff (1734-1794) observe l’épigenèse au microscope sur l’embryon de Poulet et qu’il ébauche une explication de l’embryogenèse (développement de l’embryon) à partir de trois feuillets fondamentaux concentriques : l’ectoderme (feuillet externe), le mésoderme (feuillet moyen) et l’endoderme (feuillet interne). Pourtant, sur le fond, le problème n’est pas vraiment résolu : l’épigenèse révélée par le microscope optique pourrait cacher une préformation située à l’échelon inframicroscopique, inaccessible pour le moment.
Buffon, l’as du compromis, va donc défendre une épigenèse relative, admettant une part de préformation, mais refusant l’idée des germes préexistants et emboîtés. Car on ignore en effet tout de la nature des « matières sexuelles », et de façon plus générale de l’organisation infra-anatomique des êtres vivants, sans parler de leur composition chimique.
Les germes version Buffon
Nous avons vu (dans cette revue Buffon : le moule intérieur dans la nutrition) que Buffon avait adjoint à son modèle du moule intérieur la notion de germe. Ce terme est employé alors dans des sens différents, ce qui rend confus le discours de Buffon. Or il a travaillé en 1748 avec l’abbé anglais Needham (1713-1781), lorsque celui-ci est de passage à Paris. Rompu aux techniques microscopiques, il étudie d’étranges et très petits animaux aquatiques, les Hydres, à l’étonnante propriété de régénération à partir d’un tout petit fragment. En fait, Needham distingue le cas des animaux inférieurs où, comme chez l’Hydre, n’importe quel « germe » (très petit morceau) constitutif de l’animal peut régénérer un animal entier, par opposition aux organismes supérieurs où cette régénération est impossible, ou rarement très partielle (ex. queue de certains lézards). Chez l’Hydre, le « germe » constitutif, supposé mais non identifié, peut redonner, une fois isolé, un animal entier. C’est une forme de clonage qui ressemble au bouturage chez les plantes. Si Buffon reste incapable d’expliquer pourquoi le phénomène ne se produit pas chez les êtres supérieurs, il adopte à cette occasion une théorie de l’organisation du vivant très audacieuse et en avance sur son temps, quoique inspirée du modèle du médecin et chimiste allemand Stahl (1659-1734).
Trois niveaux d’organisation emboîtés
Buffon conçoit l’existence de trois niveaux emboîtés. Or les organes sont composés de tissus dont on a, à l’époque, une perception très rudimentaire. La thèse qui domine l’histologie (étude des tissus) est celle de la « fibre » : les tissus seraient formés de fibres. Le muscle donne une vague idée de ce que cela peut être, quand on observe un morceau de viande. Pour ajouter à la confusion, on emploie aussi le mot cellule, mais dans un sens purement descriptif et pour désigner des espaces, des « vides », autrement dit des lacunes entre les fibres.
Pourtant, le microscope de l’époque offre déjà des agrandissements permettant de voir les cellules de la biologie moderne. Mais personne ne comprend ce que l’on voit sous l’oculaire, sans compter que l’on sait assez mal préparer les tissus pour l’observation microscopique. Ainsi, lorsqu’au XVIIe siècle déjà l’anglais Hooke (1635-1703) eut observé au microscope une fine lamelle de liège, il ne put comprendre la signification des logettes ou cellules aux formes géométriques qu’il y avait vues. Et au siècle suivant, Buffon ne réussit pas mieux, qui s’obstinera à ne voir sous les lentilles que ses fameuses « molécules organiques vivantes » (voir dans cette revue Buffon : le moule intérieur dans la nutrition). Paradoxalement, cette erreur d’observation et surtout d’interprétation lui apporte de l’eau à son moulin des trois niveaux d’organisation. Le premier niveau est précisément celui de ces « molécules organiques vivantes » qui constituent la matière la plus caractéristique des êtres vivants. Ce sont elles qu’il croit avoir observées au microscope. Le deuxième niveau est celui des « germes », unités tout à fait originales et censées contenir les molécules organiques vivantes. Il s’agit d’une pure spéculation de sa part, mais il suffirait aujourd’hui de remplacer le mot germe par le mot cellule au sens actuel, et on y serait presque ! Mais n’allons pas si vite en besogne. Le troisième niveau est représenté par les tissus de l’organisme composés d’un assemblage de germes. L’architecture est séduisante et non dénuée de logique.
Pour la construire, Buffon s’est inspiré de l’analogie avec la structure des cristaux établie au siècle précédent, notamment par le danois Niels Steensen dit Sténon (1638–1686). On savait en effet depuis pas mal de temps qu’un cristal est formé par l’association de structures en tout point identiques mais beaucoup plus petites. Buffon imagine un modèle similaire pour les êtres vivants. Cette construction de l’esprit est audacieuse, mais les preuves sont maigres et le discours de Buffon est en réalité beaucoup plus confus que la présentation qui est faite ici, rétrospectivement éclairée par les connaissances actuelles.
Le schéma de la génération
Fort de cette théorie et de ses observations au microscope, Buffon propose un schéma de la reproduction, certes fortement inspiré de Descartes et que nous avons reconstitué ci-après. 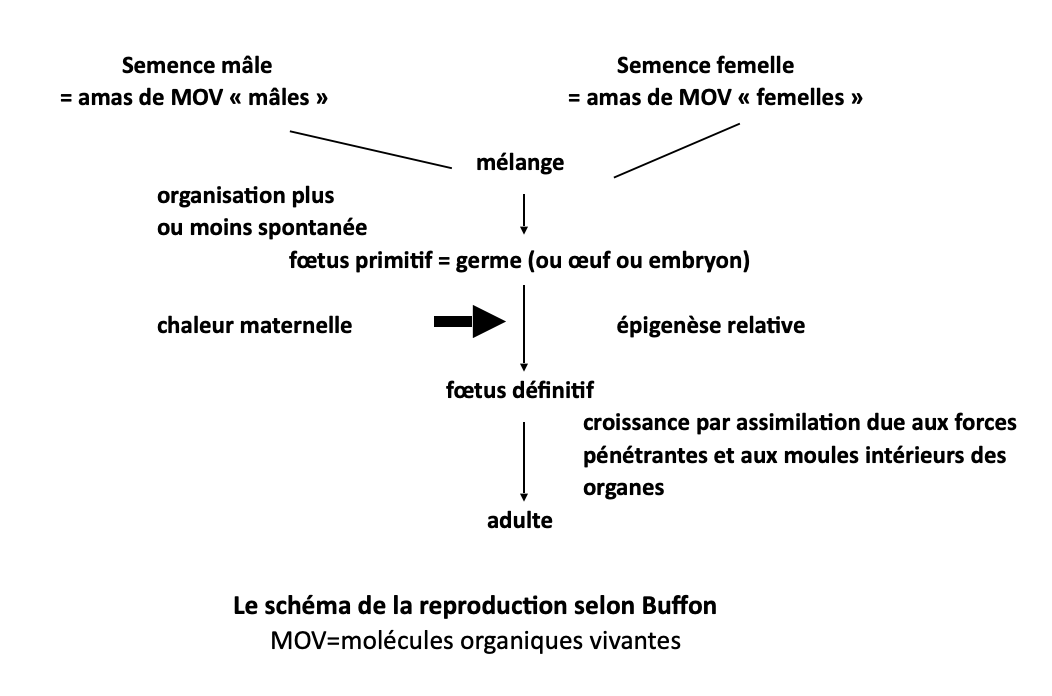
Précisons que s’il observe les « animalcules » (spermatozoïdes) dans le sperme et les follicules ovariens, il s’obstine à les considérer comme ses fameuses « molécules organiques vivantes », matériaux nécessairement « mécanistes » dont le mélange produit un germe ou embryon primitif. Inorganisé, il se développera en faisant apparaitre les ébauches des organes sous l’effet de la chaleur maternelle : la survivance aristotélicienne et cartésienne est patente.
Il ne lui reste plus qu’à y ajouter le fameux moule intérieur, dont il existe selon lui autant de variantes que d’organes. Car il faut bien expliquer – c’est là l’originalité du schéma buffonien – comment les matériaux sont assemblés de façon particulière pour former qui du foie, de la rate, des muscles ou du cerveau. Les moules intérieurs des organes contiennent – certes sans qu’il puisse en donner un quelconque support matériel – l’information qui va prescrire l’assemblage et l’organisation spécifique de ces matériaux d’origine alimentaire dans chacun des organes. Or un autre problème demeure non résolu à cette époque : l’origine et les mécanismes de l’hérédité.
Le moule intérieur et l’hérédité
Le phénomène lui-même ne fait pas de doute, mais comment rendre compte du fait que « les chiens ne font pas des chats » et que les enfants ressemblent davantage, globalement, à leurs parents qu’aux autres congénères ? Les travaux du célèbre moine Gregor Mendel datent du XIXe siècle et seront demeurés très peu connus jusqu’au début du XXe siècle. Qu’à cela ne tienne, le moule intérieur arrive à la rescousse. Revoyons le schéma ci-dessus. Les semences mâles et femelles s’unissent pour former l’œuf ou germe primitif : la participation à parité des deux sexes est à ce moment assez largement admise, quoique l’abbé et naturaliste italien Spallanzani (1729-1799) continue de défendre une théorie oviste, où la semence mâle n’a qu’un rôle de déclencheur du développement de l’œuf. Sous l’influence de Mauperthuis (1698-1759), la construction spéculative de Buffon est là encore admirable : les semences mâle et femelle en tant qu’amas de « molécules organiques vivantes » doivent, par nécessité, être différentes. Il faut donc que les molécules de ces amas ne soient pas les mêmes, en tout cas pas exactement les mêmes, entre la semence mâle et la semence femelle. Or comment obtenir ce résultat ? Par l’effet des moules intérieurs des différentes organes, qui en quelque sorte prescrivent (toujours l’information) à certaines molécules de rejoindre les organes génitaux pour y former les semences. Ce faisant, les semences contiennent des ingrédients qui du père, qui de la mère, dont l’assemblage fournit à la fois un être original et ayant des traits hérités de ses parents. La boucle est bouclée.
En somme, Buffon a eu le mérite d’essayer de « tricoter » tout ce qu’on pouvait savoir sur le vivant à son époque afin de bâtir une explication qui puisse à la fois aller au-delà de simples mots et s’accorder avec les données disponibles, encore assez maigres à l’époque. Son moule intérieur aux multiples usages est un emprunt exhumé du siècle précédent. Pour ce qui est de la reproduction, il joue le rôle de mémoire intergénérationnelle. Gardons-nous toutefois de surinterpréter : il ne préfigure en aucun cas l’ADN quoiqu’il en ait la fonction d’information. N’est-ce alors qu’une simple métaphore ? Ce n’est certainement pas une théorie. Il peut néanmoins prétendre au statut de modèle. Les modèles étant, comme on sait, le plus souvent analogiques en biologie.
Références bibliographiques
Revue de l’APBG – bulletin N°4 1988, article de Bruno Alexandre.
Buffon, Histoire naturelle – Choix et préface de Jean Varloot, Folio Gallimard 1984.
Jacques Roger, Buffon, un philosophe au Jardin du Roi, Fayard 1989.
Jacques Roger, Les sciences de la vie dans la pensée française du dix-huitième siècle, Armand Colin Paris 1963.
Yves Zarka (avec la coll de Germain M-F), Buffon, le naturaliste philosophe, Chemins de tr@verse 2013.