English
English Español
Español Français
Français Deutsch
Deutsch Italiano
Italiano Português
Português
We are surrounded by so many different forms of nature, that of course we take all for granted. The yellow banana, the tall pine trees, the colourful peacock... They are what they are, and we never ask whether they might have been different: black bananas, red pine trees, a differently coloured peacock? This brings us to the general question: things as they are as a must, namely evolution had no other way... Or isn’t instead so, that evolution, constructing things as they are, took a chance pathway, and might just as well have constructed black bananas and red pine trees?
Accepting evolution in its general terms, we also accept the notion of contingency. This means, that evolution proceeds without a predetermined plan – there was no plan to make the yellow banana, it came this way because of the vagaries of the evolution pathway – in a figurative way, a structure grows in evolution according to the changes in environment, for example it goes right because there it acquires more stability, then turns left because of a change of climate, then again right because of a lowering of acidity... Seen this way, it appears than clear that the banana might have turned out to be black... or it might have never been born. And if this is true for the banana, why not also for the human kind? Might have been possible that homo might have not be formed by the vagaries of evolution?
Thus, Stephen Gould, one of the most authoritative defendants of contingency, has to write about the Cambrian revolution, which caused the formation of multi-cellular organism on Earth after 2 billion years of dominance of unicellular organisms: «Run the tape again, and the first step from prokariots to eukatiots may have taken 12 billions years instead of 2» implying that a change in one of the variables, (temperature, pH, salinity, pressure, concentrations…) might had prevented the Cambrian revolution then. And in this case, simply, mankind would not be here.
Here we would open a big debate, which would also involve religion and the principles of determinism. In fact, the controversy between contingency and determinism is a classic in the field of life science and philosophy.
Determinism in its broadest meaning requires that life on Earth be the product of a series of subsequent steps linked with each other by causal relation, all in key with the natural laws. In this way, the origin of life on our planet would be an inescapable outcome once that the “right” initial conditions were established. And from this vision comes the assumption that mankind, and its attributes, such as intelligence and consciousness, are inevitable consequences of the origin of life.
The advent of synthetic biology
Is there a way to tackle the controversy determinism/contingency from an experimental point of view?
In what is called the chemical synthetic biology, modern chemists and molecular biologists are capable of constructing in the laboratory structures of life which were not formed by evolution – and ask, what would had been the consequences if evolution would have gone that way. For example: nucleic acids, all of them in our life, possess ribose (a cyclic sugar structure with five carbon atoms). Why not glucose, the normal sugar, much more common and more stable, which is so widespread in nature? Well, it is possible to synthetize nucleic acids having glucose instead of ribose – and show that the corresponding macromolecules would not have had the properties of building double helical structures capable of self-reproduction... (see the work of Eschenmoser and co-workers at the ETH Zurich).
Several examples of this kind could be nowadays given; they are part of our biochemical literature. I would like in what follow, to give an example based on “our” proteins, namely the proteins which make our life on Earth.
The never born proteins
There are about 30,000 different proteins in our body, and if we consider the slight variations of our proteins in all the other living organisms – from mammals to insects to plants to microorganisms – (e.g., look at haemoglobin, chymotrypsin, wool, silk, tissue proteins, cheratins etc), we should arrive at two orders of magnitude higher. And there are many organisms, mostly at the level of micro-organisms, which we have not yet investigated. So, let’s be generous, and say that we may have on Earth something like one billion (109) different proteins.
Indeed, an impressive number – at the same time ridiculously small compared to the theoretical number of possible proteins. Let’s see that.
Consider a protein chain length of 100 residues, and ask the question: in how many way can one, in principle, realize such a structure, considering that we have at our disposal, in each of the 100 positions of the chain, twenty different amino acid structures?
Answer: the number of mathematically possible different chains is 20100. Probably an extremely large fraction of them are energetically impossible, but even if only one in ten billions would be energetically allowed, the resulting number would be staggering, say 2090. This is a ten followed by 270 zeroes, a number so large that does not make any sense from a practical point of view.
There are occasionally claims that the number of proteins actually tested in the course of evolution, are very many. But cannot be all, and such a consideration is however irrelevant for the sake of the present argument: even if an immensely larger number of proteins would have been tested, the fact is, that our extant proteins are very few.
Then, how and why have those few been selected, how and why all others have been discarded? Determinism or contingency?
Just to pin down a figure about all this, let us consider our two extreme figures, 2090 and 1014. The ratio between these two figures would correspond (within a few orders of magnitude) to the ratio between the whole sand of the Sahara Desert and one single grain of sand (main photo).
We humans thrive on a grain of sand. And all other grains of sand in the vast and deep Sahara represent sets of “never born proteins” (NBP), proteins that are not with us presently. The Sahara Desert, so vast and several kilometres deep, represents an immense reservoir of proteins which in principle await to be conceived and waken up. But aside from that, the first most stringent question arising from these simple-minded considerations is: how then has that grain of sand – our few proteins on Earth – been selected out of the immensity of possibilities?
Talking about particular properties, let us focus on folding, as this is the property that more than any other epitomizes the functionality of globular proteins: is folding one specific property of our globular proteins, or is something that any other non-born proteins can share?
Let us test this experimentally: let us make a different grain of sand and see how the corresponding proteins look like – in particular let us measure their frequency of folding within a library of “never born proteins” (NBP), implying by this name that they are not in our data bank and that they are not designed on the basis of sequences chosen by evolution.
To make NBP is conceptually easy by DNA recombinant techniques: if you ask your computer to make a totally random DNA sequence, say 150 residues long, the probability that by doing so you would hit an extant gene (and by inference an extant 50 residue long protein) is as small as that ratio between a grain of sand and the whole Sahara Desert.
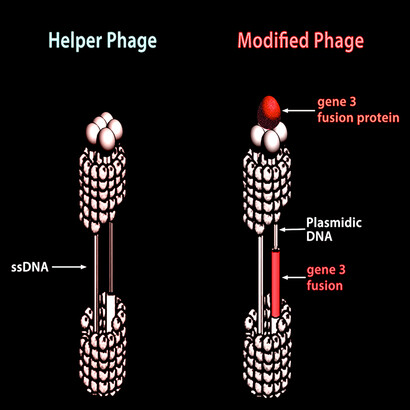
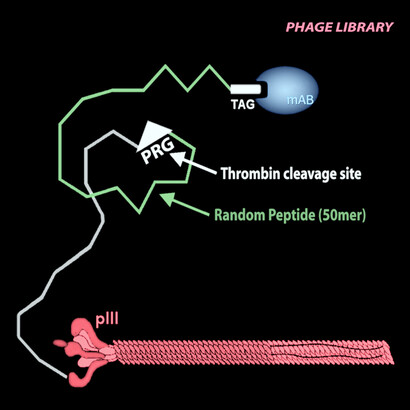
So, let us make a library of say 1013 such random DNA sequences, let us insert them –each by one – in the genome of a phage; and let us multiply these phage in an E. coli host (this is the well known phage display technique, see Figures 1 and 3).
This has been done (see paper by Chiarabelli et al. in the references), and in fact large libraries of 50 residues long proteins have been expressed in this way. A limited but statistically significant number of clones has been randomly selected and investigated, and none of the corresponding 50 residues long chains was in the data bank, not even under the most stringent criteria of similarity.
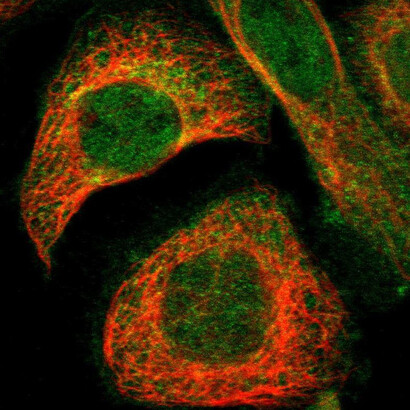
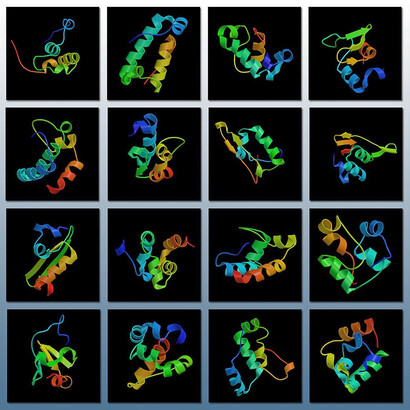
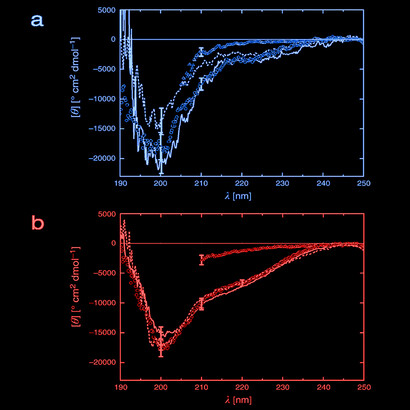
The three-dimensional structure of these totally random de novo NBP proteins has been predicted theoretically, and the results are shown in the Figure 5. At first sight, these NBP look very similar to those which we already know: the familiar helical stretches, alternated by β-structures, gamma turns and aperiodic loops. A couple of these NBP have been studied spectroscopically, by circular dichroism, as shown in Figures 2 and 6.
The CD spectra are consistent with a folding with a significant helical structure, and in both cases the folding is thermo-reversible, as judged by the classic guanidinium chloride denaturation-renaturation test. All these lines of evidence appear to suggest that “our” globular proteins are not exceptional at all from the point of view of folding, or water solubility, or thermodynamic globular stability.
And then let us go back to the question determinism versus contingency. It is tempting to use the above data to say that “our” proteins have not been selected because of structural prerequisites such as folding, water solubility or thermodynamic stability. Then, the most likely interpretation is a plead for contingency, namely our proteins are the way they are due to the accidents of contingency during their chemical evolution; and then, being fabricated in this way, they happened to foster life.
However, the most important “meta-message” coming from all that is represented in the main photo. Our proteins, namely our life, are just a grain of sand in the Sahara Desert. But then, our Sahara Desert, our globe, is just a grain of sand in the planetary system; and this is just a grain of sand in our galaxy; and our galaxy is just a grain of sand in our universe. And, where is mankind life in all this?
I remember when, still a child, I was watching television, and my eyes fell on an ant which was climbing up the screen. Would ever that ant become aware of the functioning of the TV? Isn’t the same with man climbing up into the space with his little ship (Figure 2)?
Article written in collaboration with Prof. Angelo Merante
References
Chiarabelli, C., and Luisi, P. L. (2014). Chemical synthetic biology. Science Progress, Volume 97, pp. 48–61.
Chiarabelli, C., Vrijbloed, J. W., Thomas, R. M., and Luisi, P. L. (2006a). Investigation of de novo totally random biosequences, Part I: Chemistry and Biodiversity, 3, 827–839.
Chiarabelli, C., Vrijbloed, J. W, De Lucrezia, D., Thomas, R. M., Stano, P., Polticelli, F., Ottone, T., Papa, E., Luisi, P. L. (2006b). Investigation of de novo totally random biosequences, Part II: Chemistry and Biodiversity, 3, 840–859.
De Duve, C. (2002). Life Evolving: Molecules, Mind and Meaning. Oxford University Press.
Luisi, P.L. The emergence of life, Cambridge University Press – Second Edition, 2016.
Gould, S. J. (1989). Wonderful Life. Penguin Books.