English
English Español
Español Français
Français Deutsch
Deutsch Italiano
Italiano Português
Português
Starting with a disclaimer
When the subject is self-replication, in the general context of life and nature, we think immediately to DNA. In fact, this is the driving force for the eternal flow from one generation to the other – being roses or elephants or microbes – and also, due to perturbations and mistakes in the replication mechanism itself, to the formation of new forms of life and new species – till the tree of life we know today.
The disclaimer, as we mentioned in the starting title of this first section, is important, and due to the fact that most common readers, but also simple-minded scientists, think that DNA is really able to self-replicate alone. DNA alone does not do anything. If you have DNA in a solution and provide all possible low molecular weight nutrients, salts, mononucleotides (the monomers of the nucleic acids) – nothing happens, there is no reproduction, no chemistry at all. DNA can give rise to self-reproduction only with the help of a family of enzymes, which are specific proteins which catalyze all various and several steps of the mechanism of self-replication. All self-replicating DNA of this world [1] is made by the living cells, or can even be done in vitro, but always and only with the help of that specific family of proteins – and of course in the presence of excess of nucleotides and energy bearing molecules like ATP (adenosine triphosphate). The bottom line is that DNA self-replication is a complex mechanism.
Now, all this is obvious to anyone who has followed a basic course of biochemistry, even at an undergraduate level. But in the majority of people, the idea that DNA is the only hero of all reproduction appears to be dominant in the collective thinking. This has to do with general ignorance in science, but it is also an interesting social phenomenon. As emphasized by Evelyne Fox-Keller in her well-known book, The Century of the Gene, and by several other open-mined scientists in this last period, we live in a world dominated by the notions of gene, genome, genetic engineering, a world in which DNA has become indeed the main protagonist. To the point that in many camps of the research biology – first of all the field of the origin of life – there is the unfortunate equation DNA = life. As if life would be due, or caused, by DNA alone or mostly by DNA.
Personally, I believe that this equation needs another strong disclaimer, and that actually this equation has been very detrimental to the research around the question “what is life?” and/or the complementary question “what is the origin of life?”
We are dealing with a reductionist, very restricted view, and I had the opportunity to discuss this in detail in my recent book, The Emergence of Life. And it has been emphasized in great detail that life and/or its origin cannot be ascribed to a single molecule – it is in fact a systems view, where many independent components have to interact with each other to produce a web, a wholeness, with the emergent property that we call life (see for example Capra and Luisi, 2014, or Noble, 2006).
Self-replication and self-reproduction
Not that he above disclaimers take away anything from the extraordinary beauty and molecular power of DNA, or RNA. DNA cannot reproduce by itself alone, but is the only molecular structure which has, encoded in its linear form, the information to reproduce itself; not only, as in its linear structure it has also the information of how to construct proteins – two complementary functions which are, of course, of dramatic importance for all living organisms.
Let us proceed with order, starting with a semantic note about the two terms, self-replication and self-reproduction. Although often treated synonymously, there is in fact a difference. Self-replication (the word comes from the Latin term, replica) means a faithful molecular copy, while self-reproduction refers rather to a statistical process of making very similar things. Thus, cells self-reproduce, while molecules self-replicate. It is not simply a semantic question, as according to Dyson (1985), self-reproduction processes, being less precise, may have started first in the early evolution, and preceded self-replication processes, which require more complex control and editing.
Self-replication and non-linearity
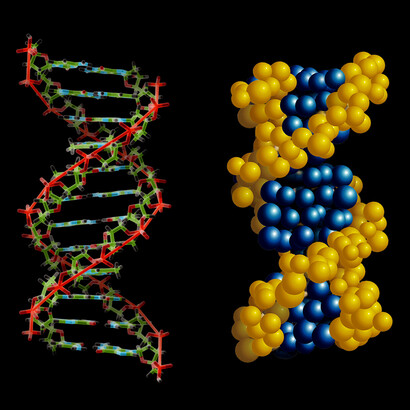
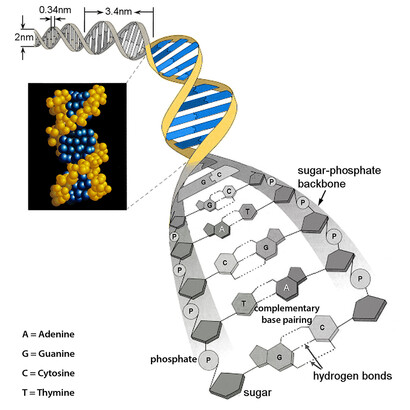
The double-strand nature of DNA (see Figure 1) was discovered by Watson and Crick in 1953, a memorable year for biology indeed. There are four different nucleotides forming DNA, the famous bases Adenine (A), guanine (G), thymine (T) and cytosine (C), whereby the two right-handed strands of DNA are kept together by the so-called complementarity of the two base pairs, A with T, C with G, as can be seen more clearly from Figure 2. This complementarity is the most ingenious device for self-replication, in the sense that each of the two strands contains the information for making a complementary one. Suppose in fact to “lose” one of the two strands – you are left with only one! But you can then immediately reconstruct the missing one, by setting a T in face of an A of your strand, then a C facing a G, and so on – this is clearly shown in Figure 2. And this happens when DNA self-replicates.
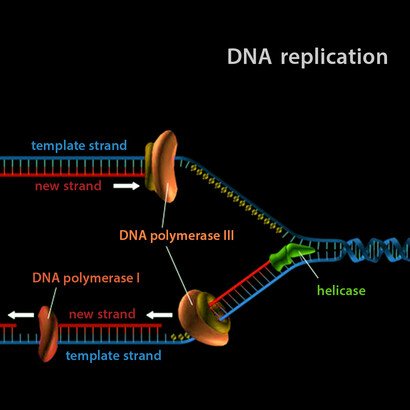
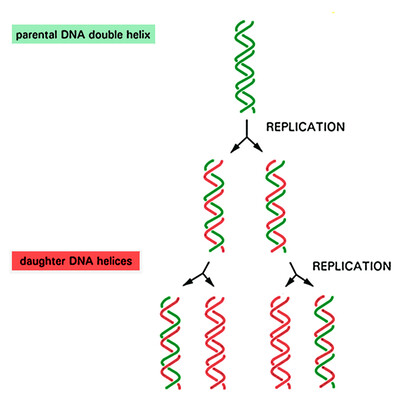
Figures 3 and 4 illustrate the well-known semi-conservative mechanism of DNA replication, which is the basis of the meiosis and mitosis processes in cell reproduction. This process, as mentioned in the text, takes place with the help of several specialized enzymatic proteins. Here, semi-conservative means that the parent double stranded molecule is divided in two, and each old strand makes a novel double strand with fresh material. So, if the parent strand would be labeled somehow, this label would be diluted by one half at each step of the duplication.
And now notice the most important feature in this process: the non-linearity. What does this mean? A linear growth process is of the type: 1, 2, 3, 4, 5… but DNA grows 1, 2, 4, 8, 16, 32… To understand the dramatic relevance of this difference, consider a chemical process which makes A→B in one second – then to make one million of B, you need one million seconds. But if you have B→2B→4B→8B→16B…, to make one million B you need only 20 seconds. That’s incredible, isn’t?
The case of self-reproduction
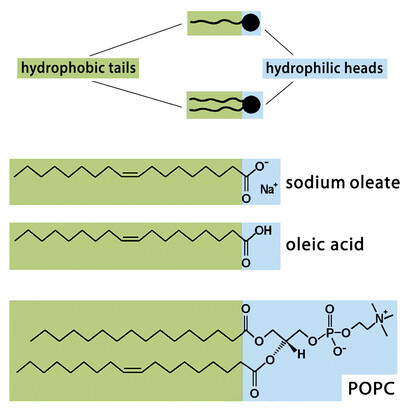
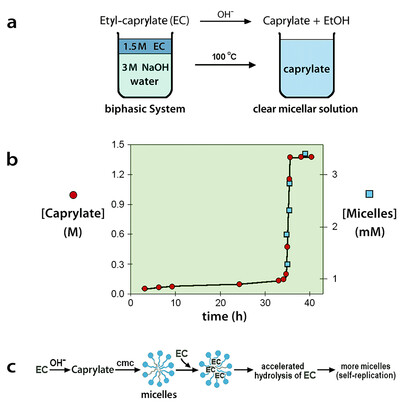
We have spoken until now of self-replication of DNA, which is a linear polymer. Are there also examples in the case of models of cellular structures? The answer is yes, with micelles and vesicles. Let us recall in more detail the case of the micellar aqueous systems. The chemistry is based on fatty acids, see Figure 5, with the additional observation that fatty acids are considered possible candidates for the first prebiotic membranes. The experimental apparatus is particularly simple (see Figure 6), also a reminder of a possible prebiotic situation: the water-insoluble ethyl caprylate (a water-insoluble simple ester) is overlaid on an aqueous alkaline solution, so that at the macroscopic interphase there is a hydrolysis reaction that produces caprylate ions. The reaction is initially very slow, as shown in the Figure 6, but eventually the critical micelle concentration (namely, the concentration at which the micelles begin to form in solution) is reached, and then the caprylate micelles are rapidly formed.
Aqueous micelles can actually be seen as lipophilic spherical surfaces, and the main reason why soap takes away the grease from our hands, lies in the fact that grease is efficiently incorporated by the lipophilic core of the aqueous soap micelles. By the same mechanism, the lipophilic ethyl caprylate (EC) is avidly up-taken by the caprylate micelles. The efficient molecular dispersion of EC on the micellar surface speeds up its hydrolysis (a kind of physical micellar catalysis), and caprylate ions are rapidly formed. The result is the formation of more micelles, as illustrated in the Figure 6. However, more micelles determine more uptake of the water-insoluble EC, with the formation of more and more micelles: a typical autocatalytic behavior (Bachmann et al., 1992).
In the following years, this type of self-reproduction experiment was extended to vesicles. Let us emphasize again that the mechanism of self-reproduction of micelles and vesicles can be considered autopoietic, (see Luisi, 2016) since growth and eventually division comes from within the structure itself (in vesicles, initial reaction takes place on the bilayer, but the bilayer is part of the structure).
It is important to point out the main message of these experiments, which is the possibility of a spontaneous self-reproduction of spherical compartments. Since these can be considered as models and/or precursors of biological cells, the hypothesis was put forward, (Bachmann et al., 1992), that this autocatalytic self-reproduction process might have been of relevance for the origin of life.
Concluding
As we have emphasized, DNA does not replicate when alone, and chemists, particularly those studying the origin of life, have for several decades tried to mimic self-replication processes in the lab, without using enzymes. Two things should be said in general at this regard: that yes, chemists have succeeded to produce in the laboratory self-replication and self-reproduction processes without the use of enzymes; however, these are not processes that occur spontaneously under prebiotic conditions, they have been brought to function thanks to the use of suitable reactive groups (nucleophiles, leaving groups, and the like… we cannot go here in the details).
Thus, from the one hand the notion of self-replication, that was considered a monopoly of nature, is now into the narrow space of the chemistry lab, and this should certainly be seen as a remarkable progress. However, in general, the origin of life is for science still an unsolved question, and prebiotic self-replication is one facet of this larger problem.
References
Bachmann, P. A., Luisi, P. L., and Lang, J. (1992). Autocatalytic self-replication of micelles as models for prebiotic structures. Nature, 357, 57-9.
Capra, F., and Luisi, P. L. (2014). The Systems View of Life: A Unifying Vision. Cambridge University Press.
Dyson, F. J. (1985). Origins of Life. Cambridge University Press.
Fox-Keller, E. (2002). The century of the gene, Harvard University Press.
Landenmark, H. K. E., Forgan, D. H., and Cockell, C. S. (2015). An Estimate of the Total DNA in the Biosphere. PLoS Biol, 13(6): e1002168. https://doi.org/10.1371/journal.pbio.1002168.
Luisi, P. L. (2016). The Emergence of Life: From Chemical Origins to Synthetic Biology. 2nd Edn., Cambridge University Press.
Noble, D. (2006). The Music of Life. Oxford University Press.
Von Neumann, J., edited and completed by Burks, A. W. (1966). Theory of Self-Reproducing Automata, University of Illinois Press.
[1] By the way, have you attempted to calculate how much DNA we have in our world? What do you think? A few grams… or perhaps billions of tons?
Let us make a calculation, considering that each human body consists of 15-30 trillion cells, and that each cell (except for mature erythrocytes) contains ca. 6 pg (means pico-gram) of DNA. Now consider all people living on the Earth – but also all other species, including plants, bacteria and DNA viruses – a biomass which is approximately 2 × 1012 tons.
How much DNA, then?
Well, the research group directed by Charles Cockell has calculated that the total DNA existing in the biosphere is ca. 50 billions of tons. Take account that more than 98% of this weight is due to plants (68%) and bacteria (30.2%), and only in a negligible way to animals (0.8%) and other living organisms (Landenmark et al., 2015).